马雪梅教授:氢气生物学机理(3)氢化酶
法国化学家拉瓦锡Antoine Lavoisier(1743-1794):呼吸就是一种缓慢的对碳和氢的燃烧作用,这完全类似在一盏煤油灯和蜡烛中发生的事……因此一个人可以说,从降生到这个世界并开始呼吸开始,生命的火炬点亮了他自己,直到死亡火炬才会熄灭。

氢能源被认为是21世纪的“终极能源”,其特征是氢和氧燃烧生成水释放化学能,氢气可储存、可再生、可电可燃、零污染、零碳排,是一种清洁的二次能源。生命体是否可以利用氢气作为能源呢?答案是肯定的!生命既是物质的,也是能量的,生命意味着有机体必须维持生存、生长和遗传。能源是生命机器运转的动力之源,吸收能量并将其转变为生物功是所有生命的基本特征。生物化学反应和普通化学反应的最大区别是多数生化反应是在酶的催化下进行,反应条件温和可控。现在发现的很多生物如细菌、古细菌、蓝藻等可以产生氢气或利用氢气作为能源,这得益于它们体内存在的氢化酶(Hydrogenase)。目前,很多从事氢能源研究的学者在研究氢化酶,主要是改造微生物或藻类产生氢气,也有学者做氢化酶仿生研究,通过体外重构氢化酶活性中心从而可以脱离生物活体催化产氢,这些研究的目的主要是大量产生氢气作为一种能源供给方式。关于高等生物的细胞是否存在氢化酶活性的研究很少,尚没有动物细胞代谢氢气的报道。多项研究发现植物可以产生氢气,有报道植物的叶绿体可以产生氢气,叶绿体可能起源于具有氢代谢功能的藻类,它能产氢是可以理解的,但并没有关于植物线粒体和质膜代谢氢气的研究。我们在这方面的一些探索性研究会在后边的文中阐述。既然“氢气是推动真核生物进化的关键”,真核生物是由微生物进化而来,本文将从微生物氢化酶的研究及与能量代谢的关系进行分析,从而可能对真核细胞的氢代谢功能分析提供线索。
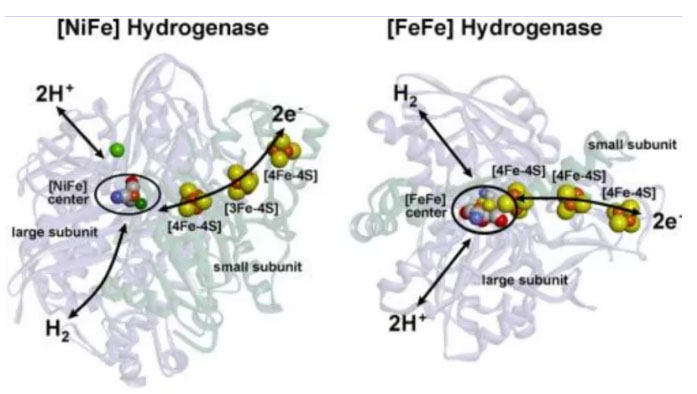
[NiFe]和[FeFe]氢化酶的结构。Lubitz Wolfgang,2014
微生物中的氢化酶
据估计,有99%的生物体具有分子氢(H2)代谢功能,这是令人惊讶的!这些物种中的大多数是微生物,包括细菌、古细菌、蓝藻和某些真核生物(如原虫)等,它们代谢H2的能力来自于体内氢化酶的表达。氢化酶是一类含有铁、镍等金属元素能够催化H2可逆氧化的金属酶。早期的地球中富含氢气(40%),可以赋予细胞利用氢气功能的氢化酶在这个时期的原始生命产生过程中被“创造”出来是合乎逻辑的。氢化酶根据活性中心所含金属组成的不同,可以分为三类,即镍铁([NiFe]),铁铁([FeFe])和唯铁([Fe])氢化酶等(Lubitz Wolfgang,2014)。
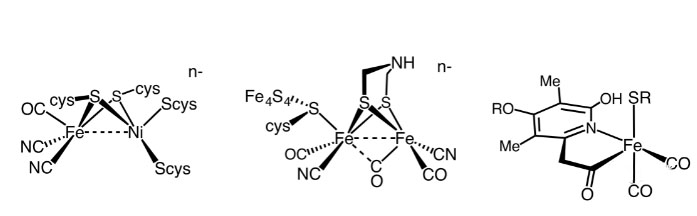
三种氢化酶活性位点的结构。By Smokefoot - Own work, CC BY-SA 4.0,
大多数[NiFe]氢化酶是吸氢型的,[FeFe]氢化酶主要产生分子氢,唯[Fe]氢化酶催化利用氢的特定反应。现有发现的氢化酶的活性中心至少含有一个铁原子,[NiFe]和[FeFe]氢化酶在结构上有一些共同的特点:每种酶都有一个活性位点和一些埋在蛋白质中的铁硫簇(Fe-S),活性中心铁原子被小的无机配体(CO和CN−)连接。唯[Fe]氢化酶只含有一个单铁活性位点,没有铁硫簇,它们只能在第二底物存在下才能活化H2。另一种分类通常是根据参与反应的氧化还原对的不同进行分类,如NAD(P)、细胞色素b型、或细胞色素c型等类型。氢化酶催化H2的可逆氧化还原反应,即氢气氧化为质子和电子,或者质子还原生成氢气的反向反应。(1) H2( ⇆ H+ + H- )⇆ 2H+ + 2e-反应发生在氢化酶的金属中心,目前比较认可的是异裂机制。在酶的金属中心H2的酸性显著增加,并导致氢分子的异裂反应,附近碱的存在极大的加速了该异裂反应。H2的产生是逆反应,涉及H+和H-的耦合。
生命的能源
氢是最轻的元素,它既能与其它元素共享电子(如C-H),也能失去电子变成质子(H+),因为氢的原子序数是 1,H+不带电子,只有由一个质子组成的原子核,是一个“赤裸的电荷”,具有很强的化学反应性。氢化酶联结了氢分子(H2)、质子(H+)和电子(e)这三个在生命能量代谢中的核心构件,想要理解氢化酶在生命系统的作用,就不得不去理解生命的能量基础。能量的转换和交换是生物体维持生存和自我繁殖的基础。活的有机体相互依存,它们从周围环境中获取能量的方式有两种:(1)它们从阳光中吸收能量;(2)它们从环境中摄取化学燃料并通过氧化这些燃料获取能量。细胞是完美的能量转换器。能够将化学的、电磁的、机械的及渗透形式的能量有效地进行相互转换。电子的流动为生命体提供能量从物质中移去电子(一或多个)叫做氧化反应,而物质接受电子(一个或多个)叫做还原反应。在生化反应中,氧化还原作用既指电子传递,又指氢原子(含一个电子和一个质子)的传递。氧化还原反应中的电子流动是活细胞中能量转化的基础。电子是一种神奇的带电粒子,电子可以共享,也可以转移,一些类型的原子容易失去(如Na+)或共享电子,而另一些原子则容易接受电子(如Cl-)。由于电子不能在溶液中存在,它们必须是原子或分子的一部分,这样,在氧化还原反应(oxidation-reduction reaction)中,同时需要电子供体和受体。这就是说,任何一个氧化反应(失去电子)的发生必须伴随一个相应的还原反应(获得电子),被氧化的物质就是电子供体,而被还原的物质就是电子受体。细胞内能量转换的核心过程是电子传递链。细胞内所有的能量转换都能追踪到电子从一种分子到另一种分子的流动,这是通过一个“下坡的”从较高电化学势到较低电化学势的电子流动,它与电路中的电子流动形式上是类似的。
生命系统某些重要的半反应在25℃和pH7下的标准还原电势

表中表示了生物体系中一些重要的氧化还原对的还原电势(E\'0/V),底部的具有较大的负值,因而具有较强的提供电子的倾向,而顶端的具有较大的正值,因而具有较强的接受电子的倾向,因此,底部右侧的物质为最强的还原剂,而顶部左侧的物质为最强的氧化剂。一般来说,电子自下而上进行传递,但是在外加能量的情况下,生物也有可以进行逆向电子传递。有如此之多的不同还原电位的氧化还原对,这就为生化反应中多种多样的电子传递链(包含多个中间电子载体)的存在提供了可能。我们现在讲的呼吸主要指有氧呼吸,也就是氧气作为末端电子受体,中间经历电子沿着由氧化还原组分组成的电子传递链的传递。呼吸作用也可以发生在没有氧的情况下,这时候硝酸盐、硫酸盐、Fe3+ 等作为末端电子受体,称之为厌氧呼吸,具体来说,称为硫酸盐呼吸、硝酸盐呼吸等。有些电子载体与膜紧密结合,有些则能自由扩散,如NAD+和NADP+(它们还是氢原子的载体)。从表中可以看出来,H2的 E\'0 负值很高,达到-0.414 V,O2 则具有较大的正值,达到+0.816 V。H2被O2氧化提供的能量是最高的。但在生物体不会直接发生这么强烈的化学反应,能量一般是在酶的催化下温和转化。ATP和被称为还原力的NADH等都是生物能的重要组成分子。氢化酶推动氢气作为生命的能源根据氢化酶的催化特性,可分为吸氢酶、放氢酶和双向氢化酶等类型,许多氢化酶是双向的,也就是说,它们既是摄取H2的酶,也是产生H2的酶,这取决于细胞中的生理环境。式(1)氢化酶催化的氧化还原反应可以分解为如下方程式:(2) H2 + Aox → 2H+ + Ared
(3) 2H+ + Dred → H2 + Dox
式(2)是氢气的氧化过程(吸氢酶活性),氧气、硝酸盐、硫酸盐、二氧化碳、富马酸等都可以作为电子受体(氧化剂,Aox);式(3)是质子还原为氢气的过程(放氢酶活性),铁氧还蛋白(ferredoxin,Fd)等作为电子供体(还原剂,Dred)。可以看出,在生物体内,小的无机或有机分子、蛋白质如硫氧还蛋白、细胞色素c3和细胞色素c6等都可以作为氢化酶的生理电子供体或者受体(Vignais, Paulette M. 2001)。氢化酶以可溶形式或膜结合形式存在于细胞周质或细胞质中;在真核细胞中,它们通常位于专门的区域。氢化酶的主要功能是通过氧化H2为生物体提供能量,并且也用于处理细胞中的多余电子,平衡细胞的氧化还原电位。生物体代表能量的分子(如NADH等)过多会产生氧化压力,氢化酶可以通过接受电子产生分子氢来去除还原当量减轻氧化压力。能量缺乏时氢化酶通过分解H2来提供电子,它们还可能参与建立代表生物势能的跨膜质子(H+)梯度。例如,真氧产碱杆菌 (Ralstonia eutropha,Re)中发现有膜结合和可溶性氢化酶。Re 的膜结合氢化酶(MBH),位于细胞膜的周质侧,具有三个亚基HoxG、K和Z,[NiFe]活性中心分解H2产生质子(H+)和电子,电子经FeS簇传递给细胞色素b(Cyt b),质子形成跨膜质子梯度推动能量货币ATP的产生。
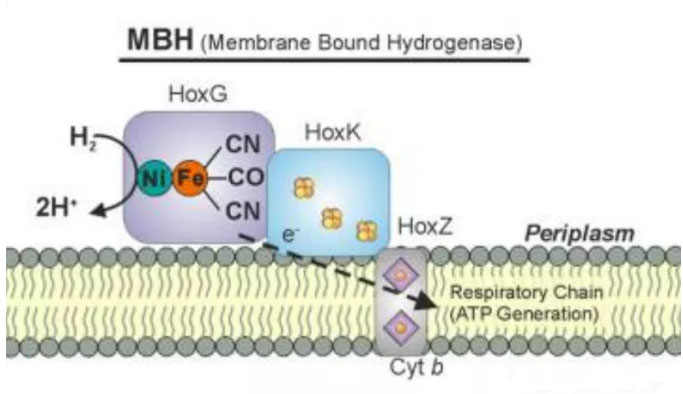
Re膜结合氢化酶。Lubitz Wolfgang,2014 Re中发现的可溶性氢化酶(SH)定位于细胞的细胞质中,SH具有四个主要亚基HoxH、U、Y和F,F也称为NAD+还原氢化酶。[NiFe]活性中心分解H2产生质子(H+)和电子,电子经FeS簇传递给NAD+产生NADH。
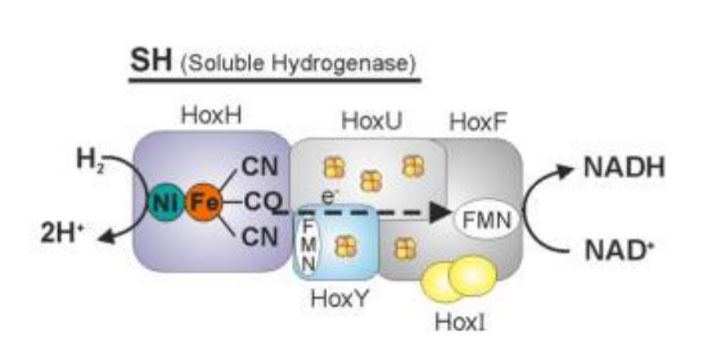
Re膜结合氢化酶。Lubitz Wolfgang,2014
氢化酶驱动的微生态系统种类繁多的微生物不仅可以通过吸氢酶利用H2作为能源,而且还具有产生H2的能力,并有可能建立由H2驱动不依赖有机碳和分子氧的生态系统。
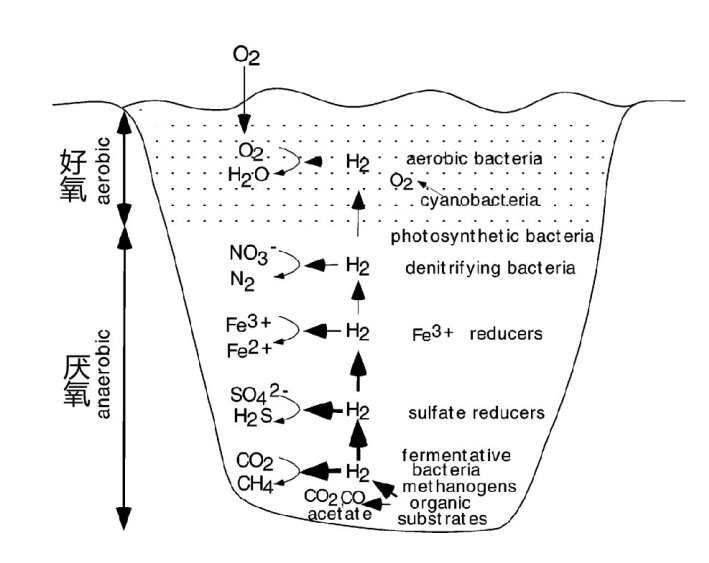
在水生分层系统中的厌氧和好氧细菌代谢。从底部往上氢气越来越少。Vignais,2007 图中显示了在湖泊、池塘等微生物生态中,能够产生或消耗H2的微生物群落催化的不同氧化还原反应的垂直分布,氧化还原电位在缺氧的底部负值更高,在与空气接触含氧气的水面附近为正值。在沉积物中,梭状芽孢杆菌通过发酵有机物释放H2和CO2。产甲烷古菌利用H2作为主要能量(还原力)还原CO2产生甲烷。硫酸盐、Fe3+、硝酸盐等电子受体的存在改变了微生物群落结构。例如脱硫弧菌,使用H2将硫酸盐还原成硫化物。某些细菌在缺氧条件下Fe3+接受电子氧化H2,还可以使用硝酸盐和氧作为电子受体。在水体表面附近,由于接触空气和蓝藻在光照下产生氧气,有氧细菌利用氧气将氢气氧化成水。厌氧和好氧环境中的H2的氧化主要是由氢化酶催化。另一个氢化酶作为能量代谢重要节点的系统是肠道微生物群落。成人肠道中聚居着大量的微生物,数量大概是人体体细胞总数的10倍,所编码的基因是人类编码基因数的150多倍。产氢和用氢的微生物在肠道内组成了庞大的氢交换市场。
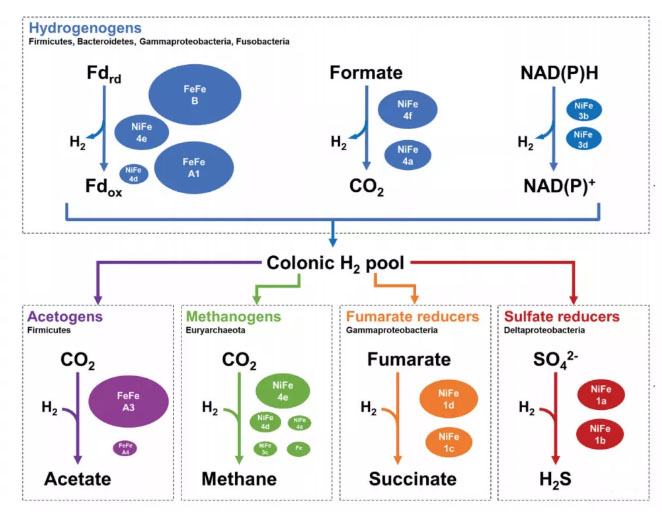
人类结肠中已知主要的H2产生和氧化途径。Patricia G. Wolf et al., 2016 微生物发酵会产生氢气,产氢微生物包括产甲烷古菌、硫酸盐还原菌(SRB)等。在产氢途径中,最主要的是[FeFe]-氢化酶催化的铁氧还蛋白(Fd)依赖的H2释放,也存在[NiFe]-氢化酶催化的NADPH或甲酸盐(Format)依赖的H2释放。产生的H2会被利用氢气为营养的微生物利用。大部分氢气是通过[FeFe]-氢化酶(A3组)与铁氧还蛋白(Fd)的呼吸耦合而被再次氧化,这是迄今为止在肠道中发现的最丰富的以氢为营养的氢化酶,能够可逆地将电子从H2传递给铁氧还蛋白和NAD,生成的还原剂可用于维持合成代谢过程、固定CO2为乙酸、甲烷,还原硫酸盐为硫化氢等。氢化酶与肠道内微生物的代谢平衡密切相关,由氢化酶介导的产氢和用氢代谢是肠道微生物群落内菌群能量代谢的重要媒介,整个肠道菌群的代谢活力和我们的一个器官的代谢活力是基本相当的,对于健康的影响巨大。摄入氢气对肠道微生物群落结构会有怎样的影响,又是如何通过肠道微生物影响健康的,这方面的内容值得深入研究。结语氢化酶的家族非常庞大。既然99%的生物体有氢化酶的活性,真核生物是由细菌和古细菌进化而来,真核生物中是否有氢化酶的类似结构?答案是肯定的,线粒体复合物 I 与氢化酶高度同源。我们研究发现植物的线粒体可能具有产生氢气的能力,这是第一个高等生物线粒体具有氢代谢能力的研究,这为我们在高等真核生物中寻找氢化酶多了点信心。
参考文献
Lubitz W, Ogata H, Rüdiger O, Reijerse E. Hydrogenases. Chem Rev. 2014 Apr 23;114(8):4081-148. doi: 10.1021/cr4005814. Epub 2014 Mar 21. PMID: 24655035.
Vignais PM, Billoud B. Occurrence, classification, and biological function of hydrogenases: an overview. Chem Rev. 2007 Oct;107(10):4206-72. doi: 10.1021/cr050196r. PMID: 17927159.
Wolf PG, Biswas A, Morales SE, Greening C, Gaskins HR. H2 metabolism is widespread and diverse among human colonic microbes. Gut Microbes. 2016 May 3;7(3):235-45. doi: 10.1080/19490976.2016.1182288. PMID: 27123663; PMCID: PMC4939926.
Vignais PM, Billoud B, Meyer J. Classification and phylogeny of hydrogenases. FEMS Microbiol Rev. 2001 Aug;25(4):455-501. doi: 10.1111/j.1574-6976.2001.tb00587.x. PMID: 11524134.






