能消化氢气的土壤细菌是好菌!
豆科作物根际有能改良土壤或提高肥力的作用,有一种观点认为是因为豆科植物根系具有产生氢气的能力,从而滋养着一批能消化氢气的细菌,这种细菌就是氢氧化菌,就是利用氧化氢气获得营养生存的细菌,这类细菌是土壤是否肥沃的重要基础。有一篇2012年的综述文章,对这个问题进行了比较全面的总结。
王瑾, 王喆之, 董忠民. 土壤氢氧化细菌促进作物生长机理研究进展[J]. 应用与环境生物学报, 2012, 18(5):853-861.

摘要:本文综述了近十年来国内外关于豆科作物根际土壤促生菌中氢氧化细菌的研究进展,讨论了根瘤释放氢气促进作物生长的可能机制.与豆科作物进行轮作、间作是提高土壤肥力、增加作物产量的一项传统的农业耕作方式.对于这种耕作方式优势的机制研究,过去大多数主要集中在土壤氮(N)元素含量的提高.而近期研究表明土壤氢氧化细菌以豆科作物根瘤菌在固氮过程中释放的氢气为能量来源进行化能自养改变土壤微生物种群结构.一些土壤氢氧化细菌通过产生1-氨基环丙烷-1-羧酸脱氨酶和根瘤菌毒素抑制植物体内乙烯的合成,促进作物生长.此外,本文进一步讨论了利用现代分子生物学方法研究氢氧化细菌促进作物生长的途径.
土壤是农业生产系统中不可缺少的基础,它不仅为农作物生长提供基本环境,维持作物产量,还影响到环境质量以及关系到动植物甚至人类的生存[1]. 二战以后,为了应对急剧增加的人口压力,各国在现代农业中利用大规模单一作物耕作而带来一场绿色革命. 降低农家肥应用量,同时过量使用化学性氮肥最终导致土壤肥力逐年直线下降. 传统意义上,土壤肥力主要来源于可测定的土壤中氮(N)、磷(P)、钾(K)等无机元素的含量. 而现代意义上,土壤的物理、化学和生物性质的改变也决定着土壤肥力的高低. 相当长的时间内土壤的质量或健康状况都决定着农业的可持续发展[2]. 为保证现代农业的可持续发展就是要提高土壤的可持续利用,而如何改善土壤耕种环境就是摆在人们面前的一个至关重要的课题.豆科(Leguminosae)作物作为可食用的高营养健康农作物,是人类食品中淀粉、蛋白质、油脂的重要来源之一,经济价值仅次于禾本科. 更重要的是豆科作物与其根瘤固氮菌互利共生作用,不仅能够将大气中的氮气转化为其自身所需要的氮肥,还能提高土壤中N元素含量而被广泛应用为饲料、绿肥和大地的覆盖物等. 大量研究和实践表明,将豆科作物与其他作物进行轮作、间作可明显改善土壤质量提高土壤肥力,并能促进下一茬农作物生长,提高产量. 同时这种耕作方式的优势与土壤中具有促生作用的微生物活动密切相关[3]. 其中,氢氧化细菌作为土壤中一大类群发挥着重要作用. 本文从微观方面综述了豆科作物根际土壤中氢氧化细菌促生机理的研究进展,以期进一步丰富自古以来采用轮作、间作以达到增产目的生物学解释,而且也为育种学家筛选优良菌株提供一定理论基础. 1 轮作、间作发展及优势轮作是一种在同一田地上有顺序的轮换种植不同作物或轮换采用不同复种方式的种植方式[4],而间作是指在同一田地上,生长季节相近或相似的两种或两种以上作物按一定比例分行或分带种植[5],二者都是中国农业生产的传统栽培方法. 早在2 000多年前的汉朝,轮作与间作已有萌芽,并在魏晋南北朝时期得到广泛发展[6]. 二者优势相似,不仅可以通过不同作物根系的形态组成差异改善土壤结构[7~9],增加土壤蓄水量及缓冲能力[9],有利于作物对营养的吸收利用[10].此外,实施轮作、间作也能打破病虫害在作物连作中的循环[9~11],降低各类合成型杀虫剂对土壤和农作物造成双重损害,起到生态防治的作用等优点. 尤其是与豆科作物轮作、间作的复种方式已经广泛地用于粮田和菜田上. 非豆科作物与豆科作物进行轮作、间作不仅能促进非豆科作物增加产量,提高单位面积土地经济收益[12~13],保证粮食安全环保,而且由于豆科作物根瘤菌的固氮作用,使土壤中N元素含量增加[7~9],进而减少化学性氮肥如铵态、硝态和酰胺态氮肥的大规模使用,节省能源. 轮作、间作制度在包括欧洲各国、美国、加拿大、澳大利亚等发达国家在内的世界各国被广泛采用[13~17].随着豆科作物与其根部的根瘤菌共生固氮作用的发现,一直以来许多研究者认为这种与豆科作物的轮作、间作的效益主要是来自豆科作物根部的残留氮的作用. 而最近的研究表明:非豆类作物的增产中只有大约25%可归结于改善后土壤中残留的氮肥营养,而大部分增产作用还不能用已有的理论解释[13, 18~21]. Dong等自2 001年以来通过大量的实验和理论推论计算,提出了一个新的氢肥理论[22]. 理论认为不含吸氢酶的根瘤菌在固氮过程中释放的氢气可促进其根际植物促生细菌(Plant Growth-promoting Rhizobecteria,PGPR),尤其是氢氧化细菌的群落生长,进一步促进作物生长.2 豆科作物根瘤菌与氢气2.1 固氮过程中氢气产生关于氢气的产生与固氮关系的研究始于上世纪40年代初.Wilso等首先报道固氮菌在固氮过程中通过固氮酶催化作用将电子传递给质子产生氢气,即每固定1个N2分子至少产生1个氢气分子[23]. 而在氮底物较低时,或者说固氮酶处于“氮饥饿”的条件下,会释放更多的氢气分子,释放积累的还原力和ATP [24]. (至今为止,生物产氢仍然是这种释放还原力的观点)
研究表明,在大田条件下,豆科作物每还原1个 N2分子,固氮酶产生大约1.5个氢气分子,因此氢气是固氮酶催化的固氮反应的副产物,消耗固氮反应33%的能量[25]. 计算表明每公顷大豆在结果期间每天可释放超过5 000 L的氢气,而固氮反应过程中用于氢气生产的能量大约为植物净光合作用的5%~7% [26]._ 豆科作物的共生固氮菌依据是否具有氢酶(uptakehydrogenase,HUP)活性可以分为两大类:一类是具有氢酶活性的固氮菌(HUP+),它可以回收利用产生的氢气并从中获得能量;另一类是缺乏氢酶活性(HUP-)的固氮菌,使固氮反应中产生的的氢气未经吸收直接扩散至根瘤表面后进入土壤. 有研究显示HUP+是固氮效率更高的固氮菌[27].
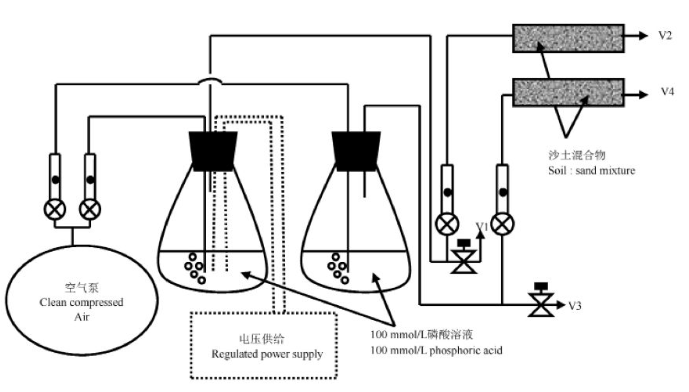
图1氢气处理装置
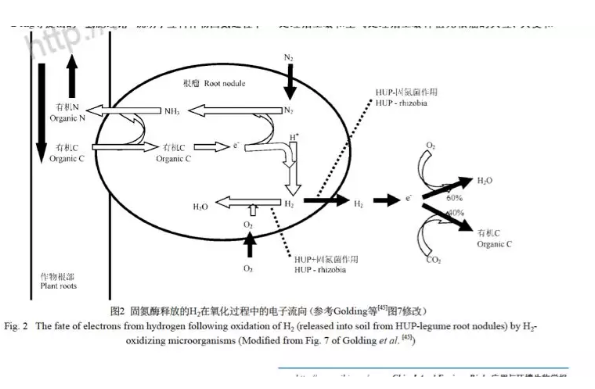
HUP体系保证了固氮酶在过量氧气和氢气抑制浓度下的活性,在氢气回收的过程中传递电子生成ATP,保证了因释放氢气而散失的能量得到回流[28~29].Conrad等对土壤中氢气的氧化做了深入的研究,得知土壤是氢氧化的主要场所,证明土壤中氢氧化细菌的存在,表明土壤中氢气被氢氧化细菌吸收利用,因此根瘤释放的氢气与土壤中细菌接触后会迅速发生氧化反应[30~32]. 同时对氢气氧化酶化学动力学(Km和Vmax)做了相关研究. Dong等在实验室模拟大田土壤氧化氢气的过程,采用持续通氢气装置进行氢气处理(图1)[26]. 其原理是利用直流电控制两根惰性电极电解水产生氢气,电流量的大小决定了阴极氢气的产量. 将产生的氢气与一定流速的空气混合后可得到已知浓度的氢气,将混合后的气体通入装有土壤的试管中,测量了16 d内土壤对氢气的吸收率,结果表明在前7 d中氢气吸收率增长缓慢,但从d 8开始吸收率突然增大,直至d 13逐渐保持水平. 此吸收曲线类似于微生物生长曲线. 在对土壤进行3周的氢气处理时,他们同时还检测了氢气、氧气和二氧化碳三者的浓度的变化,根据检测到土壤对氢气、氧气和二氧化碳的吸收数值计算出来自氢气的电子约有60%被用于还原氧气最终生成水,剩余的40%的电子则被用于还原二氧化碳最终产生有机化合物. 由此可知,释放的氢气所携带的能量中60%被氢氧化细菌作为能量而被消耗,其余的40%能量用于固定二氧化碳,提高了土壤的有机含量(图2).
3 氢气促生作用与机理研究
HUP+固氮菌较高的固氮效率不但为作物节约光合作用产生的能量,而且提高了作物自身吸收N元素的能力. 但在进化过程中固氮效率较低的HUP-固氮菌并没有被淘汰,即使是在同一物种中的进化中. 例如大豆既有与HUP+固氮菌生成的HUP+根瘤,也有与HUP-固氮菌生成的释放氢气的根瘤. 并且在现代农业生产实践中,HUP-更被育种学家及农户普遍选择使用[33]. 这表明HUP-更有助于作物生长,暗示HUP-固氮菌释放的氢气可能有益于提高农作物的产量.Dong等提出的“氢肥理论”说明了豆科作物固氮过程中产生的氢气对作物的生长具有促进作用,补充了轮作、间作获益的科学解释;氢气处理过的土壤一般可使植物茎叶或根的干重增加10%~30%或更高[22]. Dong等在温室条件下,分别用氢气处理后土壤和空气处理后土壤种植无根瘤的大豆、大麦和油菜,空气处理后土壤作为对照组,经6周后,大豆和大麦的干重与对照相比分别增加了14%和18%,种植5周的油菜干重也比对照增加了18%;另外将春小麦种植于经氢气处理30 d的土壤中,30 d后与对照相比干重提高了32%;甚至,只用5% 氢气处理的土壤与95%空气处理的土壤混合后种植春小麦,也能使春小麦的干重较对照增加21%;而且取自不同地方的土壤在经氢气处理后同样也能表现相应的促生作用[34]. Dong等分别从草坪,大田和花园中取来3种样土,经氢气处理7周后种植大麦,38 d后,大麦的干重较对照增加17%~47%;甚至仅用经氢气处理过的少量土壤接种仍具有促生作用[34]. 在大田试验中,Dong等在大麦和春小麦的每粒种子下放置3~5 mL 氢气处理过的土壤,7周后发现大麦和春小麦平均每株作物的分蘖数较对照分别增加36%和48%[34]. 由此说明,在大田条件下,混入经氢气处理过的土壤能够显著改善土壤肥力,促进作物生长. Dean等选择JH47(HUP-)和JH(HUP+)两种菌株接种大豆[35]. JH47是在JH的氢酶基因中插入Tn5转座子,使其氢酶基因沉默,释放氢气. 在大田条件下,第一年将这两种菌株分别接种大豆,经检测JH47接种大豆的土壤氢气吸收率较JH组明显增加;第二年将大麦分别种植在这两种土壤中,种植在JH47接种大豆的土壤中大麦的分蘖数、穗数以及产量也较后者显著提高.
点评:氢肥概念的本质,就是认为氢气能把土壤中依靠氢气生活的细菌养肥养多,这种细菌对提高土壤的肥沃程度,提高作物产量有很大帮助。给土壤吸入氢气固然可以,但操作起来比较困难。给土壤浇灌氢水才是最简单的方法,未来可能是农业新技术的重要方向。
氢气处理后的土壤能够促进作物的生长,已有一些研究证明是由于土壤中细菌的作用. Conrad等多年实验表明土壤中氢氧化细菌的存在,并且进一步证实土壤是氢气氧化的主要场所[31]. McLearn等研究表明,在经氢气处理过的土壤中,细菌是吸收氢气的主要部分[36]. Irvine等将36 mg/L的苯菌灵分别加入氢气处理土壤和空气处理土壤中,氢气处理土壤中春小麦干重仍显著高于后者;但使用50 mg/L链霉素和50 mg/L青霉素混合物处理两份样土后,氢气处理土壤中的春小麦干重与空气处理土壤中的春小麦干重相当[37]. 这说明土壤中有促生作用的部分是细菌而不是真菌. 由此可见,氢氧化细菌作为植物促生菌家族(PGPR)中的一大类发挥着重要的作用. 由于氢气的作用,土壤中氢氧化细菌种群发生改变,而这种变化可能就是氢气促进作物生长的原因. 因此,对植物生长根际促生菌(PGPR)中氢氧化细菌的研究将成为解释豆科作物在轮作、间作中发挥优势的一个重要方面.4 氢氧化细菌的获得及其鉴定4.1 分离及筛选对氢氧化细菌的分离,最早采用封闭系统内通入80%氢气、10% 二氧化碳和10% 氧气混合气体的配气法[38]. 这种方法局限性很多,最重要是不能很好地模拟土壤微生物生长环境而造成某些氢氧化细菌不能被分离,同时高氢气浓度也具有一定的安全隐患. Dong等通过分析大田土壤环境,推算出大田土壤中的氢气浓度在1 000×10-6以下[26]. 因此他们将氢气与空气混合以较为接近大田条件的浓度为前提,采用新的接近自然的开放系统分离氢氧化细菌[39~40],此系统能保持各种气体具有稳定浓度,达到模拟根际周围氢氧化细菌的最适生长环境的目的,可使其种群数量迅速增加. 常规细菌分离技术通常采用表明平滑的琼脂培养基,但这与土壤中的物理化学条件相差甚远. 因此,Maimaiti等在分离氢氧化细菌时将少量灭菌细土撒在平滑的培养基上,这样更利于细菌繁殖[39]. 同时,他们首次采用此装置在矿质盐固体培养基(MSA)分离出19种氢氧化细菌. 这些细菌在适当的氢气、氧气和二氧化碳浓度下以氢气作为唯一能源,二氧化碳作为碳源生长. 陈兴都等采用同样的方法也分离出20株氢氧化细菌[41]._氢氧化细菌筛选方法主要有4种[38, 42~43]:1)TTC法试验筛选氢化酶的细菌;2)氢氧化细菌的化能自养能力:3)DNA杂交试验检测氢化酶基因的存在;4)测定细菌的吸氢能力,根据氢气的减少量测定氢化酶的存在. 然而分离出的单菌落约有65%~90%都能够降低TTC的合成,显示阳性结果,但其中只有30%~65%的单菌落有氧化氢气的能力;少于30%分离的单菌落可以在有氧条件下利用氢气氧化提供能量进行自身化能自养生长;DNA杂交试验检测氢化酶基因的方法也难于检测具有氢氧化活性的细菌,因为很难判断携带氢化酶基因的细菌是否能够表达此基因并发挥功能. 因此,实践表明氧化氢能力的测定结果较可靠,其他结果可能给出假阳性的结果.4.2 基本特征及分类地位Maimaiti等分别从氢气处理土壤、大田大豆根瘤(HUP-)附近土壤以及温室大豆根瘤(HUP-)附近土壤中共分离出19株氢氧化细菌[40]. 然而由于氢氧化细菌不是一个分类学单位而是由系统学上完全不同的种类组成[43~45],它的命名来自生理上的概念,是一种不需要有机碳,在有氧的环境中氧化氢气进行无机化能营养的细菌[38, 43, 46]. Maimaiti等将它们根据Holt细菌分类手册(The Bergey’sManual of Determinative Bacteriology)分别从形态、培养状态以及生理生化特征测定等方面进行分类,结果表明,这19株菌株分别属于噬细胞菌属(Flavobacterium)、假单胞菌属(Burkholderia)和产碱菌属(Variovorax)[47]. 同时,Maimaiti等对其中一些菌种做了16SrRNA序列测定,将序列录入NCBIGenebank数据库,序列号从DQ256485到DQ256491,并与已知序列进行比对分析同源性,确定更详细的分类归属[40]. 陈兴都等从采自陕西关中地区的HUP-大豆植株根际土壤中分离出20株氢氧化细菌,并测定其基本特征[41].20株氢氧化细菌中既有革兰氏阳性菌又有革兰氏阴性菌,菌落形态小且圆,颜色分3种,乳白色、无色和淡黄色. 他们分别从19项实验测定了这20株氢氧化细菌的生理生化特性,如氧化酶活性、过氧化氢酶活性、甲基红试验、淀粉水解、纤维素水解等.4.3 群落研究由于氢氧化细菌的独特特征和大多数土壤细菌的不可培养性,土壤氢氧化细菌的分离极其困难,至今在国内外的结果仍不是很多[40, 48~49]. 然而应用现代分子生物学技术例如指纹图谱能够在土壤中探测出某些氢氧化细菌的种群变化以解释其促生作用. 目前已经有许多分子鉴定方法应用于微生物群落分析中,例如变性梯度凝胶电泳(DGGE)和温度梯度凝胶电泳(TGGE)[50]、单链构象多态性(SSCP)[51]、末端限制性片段长度多态性(T-RFLP)[44]、随机扩增多态DNA(RAPD)[52]和荧光原位杂交技术(FISH)[53]等. Zhang等应用T-RFLP技术分别分析了3种样土即实验室氢气处理土壤、温室以及大田豆科植物根际土壤中细菌的16S rRNA,得知氢气在土壤中的氧化过程改变了土壤中根际微生物的群落结构[44].Zhang等将氢气处理后的土壤细菌16SrRNA指纹变化与之前Maimaiti等分离出的19个氢氧化菌株的16S rRNA比对,但这19个单菌落的种群并没有明显变化,说明T-RFLP技术不仅检测出不同处理下土壤微生物种群结构的变化,而且能从分子基础探测新的微生物种群. Stein等使用FISH技术分析了氢气处理后土壤中微生物种群结构的变化与土壤吸收氢气和固定CO2能力的关系[53].
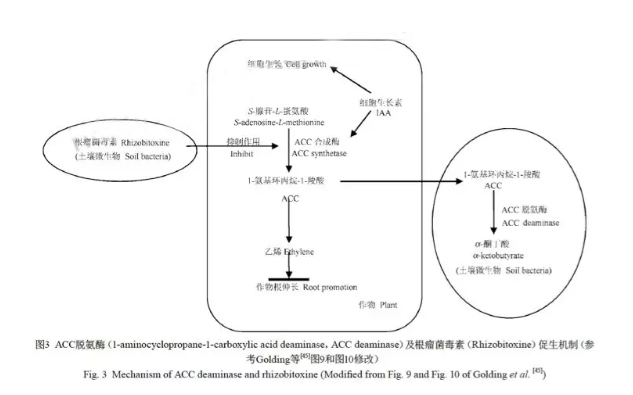
5 氢氧化细菌促生作用及机理研究Maimaiti等对分离出的19株氢氧化细菌进行春小麦幼苗促生检测,与对照相比,其中17个菌株可使春小麦种子的胚根在2 d内与对照相比伸长增加57%~250%;当用分离出的5株菌分别作用拟南芥幼苗时,其叶片数、根及茎的干重都明显较对照增加,其总体生物量增加了11%~27%[40]. 陈兴都等将分离出的20株氢氧化细菌进行小麦促生检测,结果显示其中11株可使小麦胚根与对照相比伸长增加111%~397%[41].氢氧化细菌的这种促生机制有可能是调节乙烯在植物体内合成的结果. 乙烯是作为一种重要的植物自身分泌的激素,在各个方面影响植物的生长发育[54],有研究报道在许多豆科作物中,乙烯过多会减少根瘤的产生,而根瘤的减少会影响豆科作物固氮效应[55],同时乙烯也会抑制非豆科植物根的伸长[56],使其不能更多地吸收利用土壤中的营养物质. 1-氨基环丙烷-1-羧酸(1-aminocyclopropane-1-carboxylic acid,ACC)是乙烯合成的前体(图3),许多PGPR可诱导产生ACC脱氨酶,将ACC分解为丁酮酸和氨[57],ACC水平降低导致植物体内乙烯含量的减少,从而促进幼苗的生长[40~41] . Maimaiti等已经报道6株氢氧化细菌,5株为产碱菌属(Variovorax),株为噬细胞菌属(Flavobacterium),均具有ACC脱氨酶活性[40]. 陈兴都等对分离出的11株具有明显促生能力的氢氧化菌株做了ACC分解实验,结果表明仅有1株具有ACC脱氨酶活性[41]. 此外,Hunter等报道了由豆科作物根际细菌产生的根瘤菌毒素(Rhizobitoxine)也是一种抑制乙烯生成的化学物质[55].Sugawara等研究表明根际生物素主要作用ACC合成酶,切断了ACC的合成路径,最终减少乙烯的产生(图3)[58]. Maimaiti等发现其分离的氢氧化细菌中产碱菌属(Variovorax)、噬细胞菌属(Flavobacterium)和假单胞菌属(Burkholderia)均有根瘤菌毒素的活性[40].在检测氢氧化细菌作用春小麦促生实验中,小麦的分蘖数明显增加,此种性状是受到其体内内源激素调控的结果.研究表明,细胞分裂素(Cytokinin,CTKs)在侧枝的发生和衰亡过程中起到重要的作用[59],CTKs作为第二信使参与生长素调控的休眠过程,CTKs在根部合成后运输至腋芽并在生长素(Indole Acetic Acid,IAA)的调控下影响侧芽发生[60].李春喜等研究发现不同品种间及同一品种不同种植密度间小麦分蘖节中IAA、(ZR+Z)含量有显著差异,且内源IAA、(ZR+Z)的含量尤其是二者间的比值与分蘖数的多少有密切关系[61]. 外施细胞分裂素也会增加植物侧芽的发生[62]. 已有一些PGPR已经被证实可以产生植物调节激素,如生长素、细胞分裂素和赤霉素等[63~68]. 此外,异戊烯基转移酶(Isopentenyltransferases,ipt)是细胞分裂素合成重要的限速酶,能催化异戊烯基焦磷酸和单磷酸腺苷的分解,产生作为细胞分裂素前体的异戊烯基单磷酸腺苷(Isopentenyl AMP,iAMP). 目前已被分离出的ipt酶基因大多来自微生物,编码全长为720bp,不同微生物或菌种之间ipt酶基因有一定差异,但仍具有很高的同源性. 已有一些研究证明在植物中过表达ipt酶基因可以提高CTKs含量,增加生物量,延缓植物衰老[69~70].因此,氢氧化细菌促进分蘖可能是由于部分菌株产生CTKs或产生异戊烯基转移酶而增加作物体内细胞分裂素浓度从而发挥作用.由此可见,土壤微生物与植物之间有着密不可分的关系,植物的生长影响着根际土壤微生物的种群变化,而土壤微生物分泌的活性物质也同样调节着植物的生长发育. 已有报道指出,有些豆科作物根际土壤促生菌不仅能够提高非豆科作物生物量,还能增强其自身对逆境的适应抵抗能力[71~73].
点评:氢气氧化菌如何促进土壤改良的分子机制,虽然有不少研究,但仍然不是非常清楚。不过机制虽然很重要,但效果好,环境安全才是使用的重要条件。氢气农业应用只要符合经济原理,应用不是问题。
6 展望
将豆科植物根际具有促生作用的氢氧化细菌引入农业生产,不仅能够维持土壤肥力,保证现代农业可持续发展,而且又起到增产效益,满足人类日益增长的粮食需求[74]. 然而国内外对于氢氧化细菌的相关特性研究尚未深入展开,主要由于HUP-豆科作物根际土壤中氢氧化细菌不易分离,其培养方法、种群结构分析和鉴定工作也尚存在不足,相关报道也较少. 对具有促进作物生长作用的氢氧化细菌的分离仍是菌种开发的关键. 面对土壤微生物的多样性和其生理功能的复杂性,在绝大多数土壤细菌均由于分离条件不适而不能得到有效分离的现实下,应通过分析土壤细菌生理代谢及群落结构,模拟其生存生态环境,对培养基及培养条件等做进一步改善,以便于筛选获得具有促生作用的细菌.豆科作物根际微生物促进作物生长的本质和原理还只是在猜测阶段,更多的促进作物生长的机理仍待揭示. 每1 g农田土壤中大约含有108~109个细胞,有研究推测仅细菌就约有6000~7 000种,然而其中99%以上从未被分离培养[75],因此未被研究. 利用分子生物学技术研究土壤微生物基因组是提出根瘤对土壤微生物种群结构影响的有效方法[76]. 同时,新一代高通量测序的广泛应用或许可以全面展示土壤微生物种群的表达基因[77]. 通过测定不同逆境处理下土壤中所有RNA序列(包括mRNA和非编码RNA),与Genebank比对拼接后,在获得基因序列的基础上,研究基因功能及结构,不仅能够获得土壤微生物在不同逆境胁迫下转录组的微观变化规律,而且为在RNA水平上研究豆科作物根际微生物的基因表达奠定基础. 同时也为进一步研究豆科作物根际土壤微生物对其他作物生长促生作用的机理提供分子基础,帮助育种学家筛选有利于农业生产的基因,利用根际微生物的基因表达调节促进农业作的生长,进而提高产量.