马雪梅教授谈氢气生物学机理(3):氢气是推动真核生物进化的关键
纷繁的世界中,除了我们能够看到的生命体,更多肉眼看不到的微生物构成了整个生态链的主体。目前证明,很多微生物可以产生氢气,也有很多微生物可以利用氢气作为能源。生命的起源与进化一直是人类孜孜以求尚在探索的未解之谜。包括人类在内的真核生物的细胞是如何产生的?氢气在真核细胞的进化中处于怎样的地位?对这个问题的梳理也许可以为理解氢气与生命的关系提供线索。
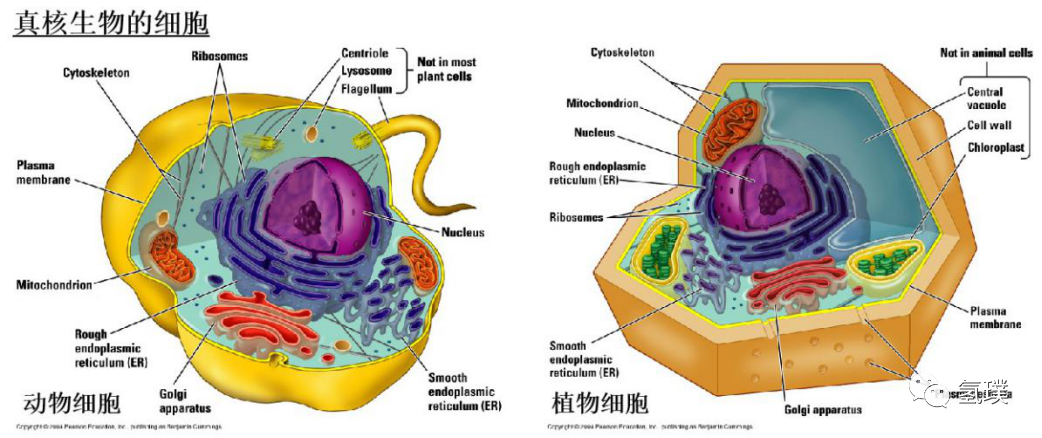
在显微镜发明之前,人们一度认为生物只有两类:植物和动物。随着显微镜观察到我们生活的环境中充满着无数肉眼不可见的微生物,生物学家把细胞生命分成了两大类:真核生物(拥有膜结构的细胞核和细胞器)和原核生物(没有细胞核和膜结构细胞器)。
20世纪70年代,古菌被发现。古菌是单细胞微生物,常生活于各种极端自然环境下,如大洋底部的高压热溢口、热泉、盐碱湖等。这些微生物与细菌有很多相似之处,即它们没有细胞核与任何其他膜结合细胞器,同时另一些特征相似于真核生物,比如基因组中存在重复序列与核小体。分子生物学的研究表明,古菌在进化上更接近真核生物,属于一种全新的生命形式。基于这些发现,卡尔·乌斯(Carl Woese)在1977年提出了细胞生命形式分类的“三域系统”,将细胞生命分为三大类:细菌域(Bacteria)、古菌域(Archaea)和真核域(Eukarya)。
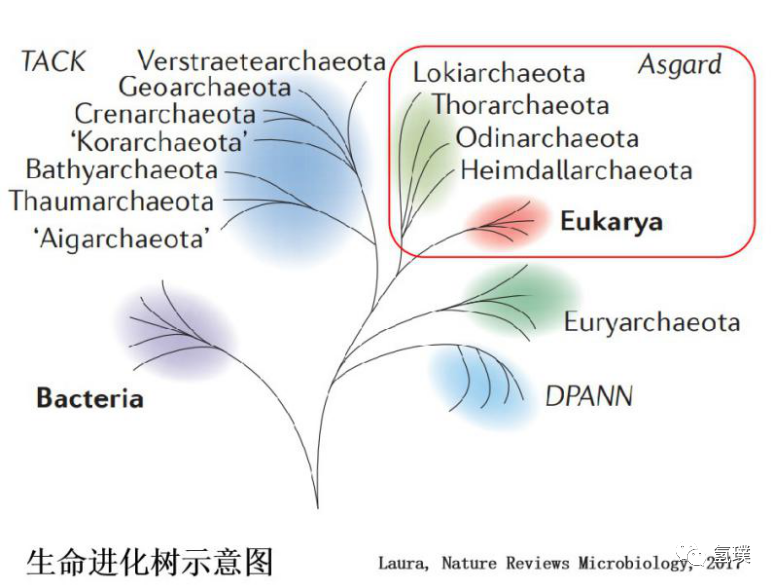
后来,Thijs Ettema 等人先后发现了4种新的古菌,分别命名为洛基、索尔、奥丁、海姆达尔,并将它们统称为阿斯加德古菌(Asgard archaea) ,随着基因组学技术被用于分类,越来越多的证据支持真核细胞与Asgard古菌在进化上密切相关。这一发现动摇了生物界分类的“三域系统”,现在认为古细菌和细菌代表了生命分类的两个主要域,真核生物是后来从这些谱系中出现,也就是新的两域分类。
地球大约在46亿年前形成,那时候地球的温度很高,从火山中喷出的气体,如水蒸气、氢气、氨、甲烷、二氧化碳、硫化氢等,构成了原始的大气层。早期地球二氧化碳浓度很高,没有氧气,同时氢气的含量高达40%,是一个适合碳基厌氧生命产生和进化的环境。
分子氧(O2)的产生对进化有深远的影响。大约 2.7-25 亿年前,蓝细菌开始利用水作为光合作用电子传输链的电子供体,产生氧气作为代谢的废物排掉,被成为大氧化事件(Great Oxidation Event,GOE)。在GOE之前,大气和海洋基本上是缺氧的,这限制了地球的生物化学循环。即使在GOE之后,氧气浓度仍保持低水平,在巴斯德点(Pasteur point)附近,也就是兼性厌氧菌从有氧代谢转变为厌氧代谢(反之亦然)时的氧分压,相当于大约 0.2 %的氧气(v/v)。尽管大气逐渐氧化,但海洋在富含硫的18亿年前元古代(Proterozic)早期真核生物产生时是缺氧的,当时海洋中的硫酸盐水平增加得很慢,限制了生物硫的循环,大气和海洋中的氧气水平仅有现在氧气水平的0.001-10%,在深海,直到5亿年前显生代(Phanerozoic)开始时仍然缺氧。这意味着真核生物前13 亿年的进化都是在低氧条件下发生的。
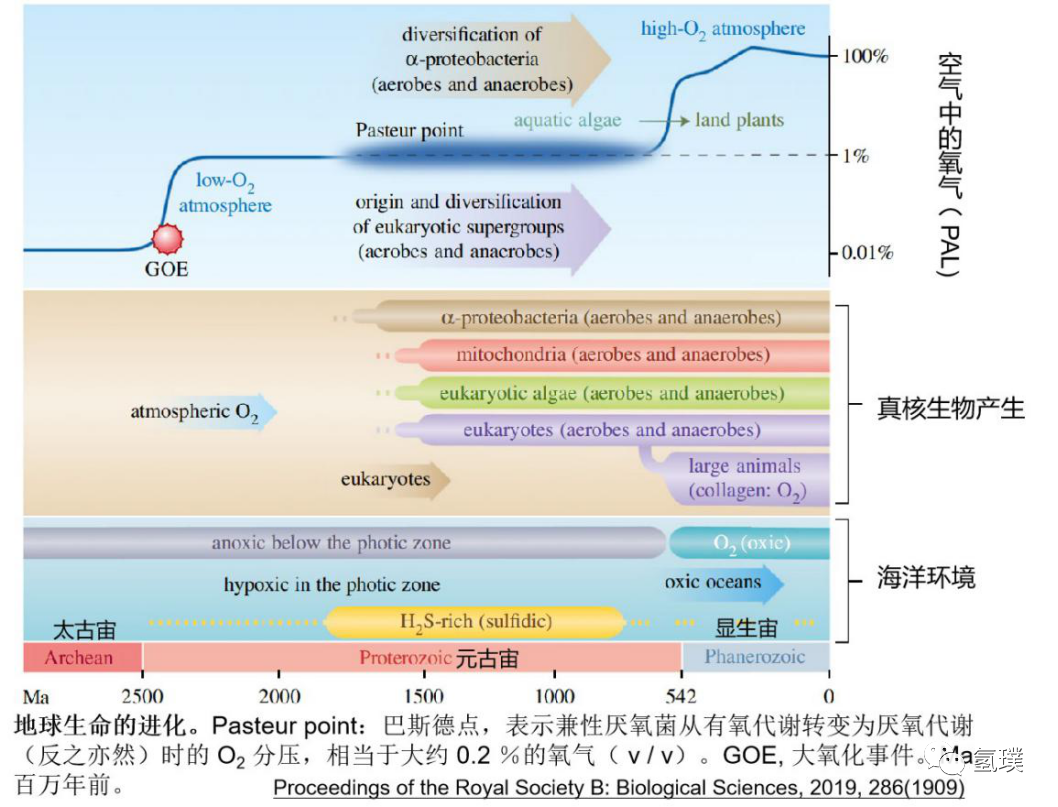
由于元古代早期的深海缺氧,真核生物有可能在浅层沉积物或光养微生物垫(phototrophic microbial mats )这类相对富氧的环境中进化。光养微生物垫可能是真核细胞发生的摇篮,它们是元古界的“森林”,微生物群落在系统发育和代谢方面丰富多样。微生物垫上层是富含氧气的蓝藻层,进行有氧光和作用,通常随后是由进行无氧光和作用的耐氧α和γ变形杆菌构成的红色层,它们合成有机物,固定在上层垫层中,这些有机物被位于更深的缺氧层中极其多样的微生物群落降解。在垂直缺氧剖面上可以区分两个宽区,其中硫酸盐还原和甲烷生成分别占主导地位。
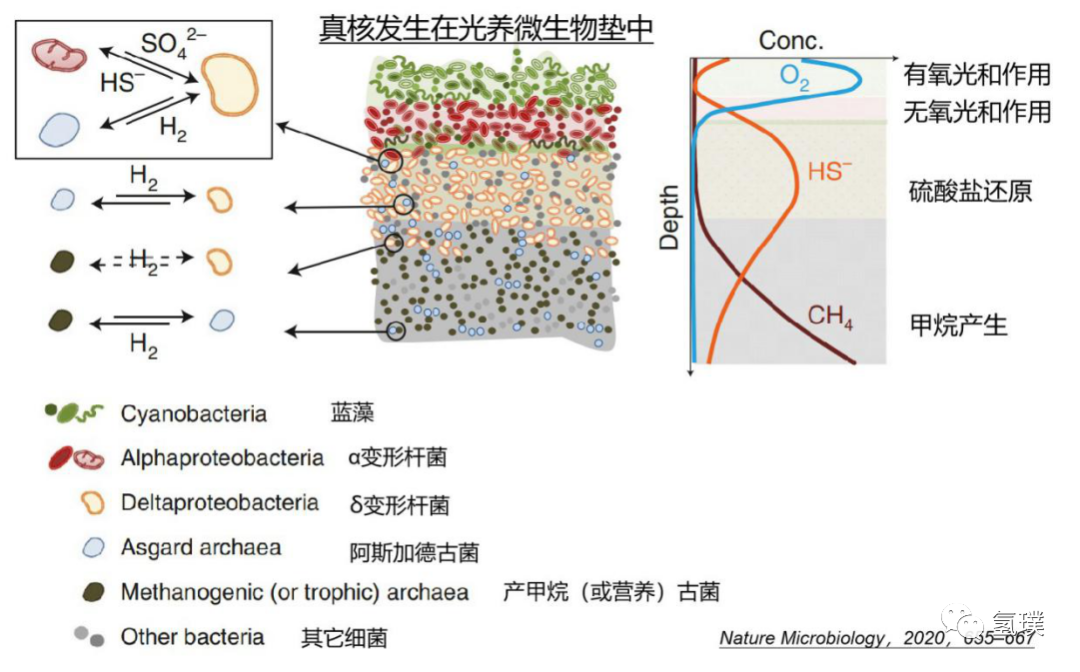
除了存在极少数无机化能细菌外,地球上生命的能量来源占绝对优势为太阳光能,而构建大分子有机化合物骨架的物质基础则是碳。对真核的自养生物(如高等植物、藻类)来说,其能量来自太阳光能,而碳源则来自大气中的CO2,但是对原核的微生物来说,情况就要复杂得多。一些微生物能够利用光作为能源(称为光能细菌),另一些能以化合物作为能源(称为化能细菌),其中以有机化合物作为能源的称为化能有机营养细菌,以无机化合物如氢气、硫化合物等作为能源的称为化能无机营养细菌。由于早期的地球环境富含氢气,产生和利用氢气的微生物丰富,微生物间氢和/或硫的转移广泛存在,微生物群落中形成互养共栖关系。
随着Asgard古菌的发现,以及微生物生态学的发展,真核细胞起源的假说也一直在完善。长期以来的观点都倾向基于古细菌和细菌之间代谢相互作用的共生模式,其中包括经典的Searcy和氢假说,以及最近的逆流(Reverse Flow)假说和共生-吞食-内生模型(Entangle-Engulf-Endogenize等。 尽管几种模型所提出的代谢相互作用上存在差异,但均比较认可的是氢气在真核细胞起源中起到核心作用。
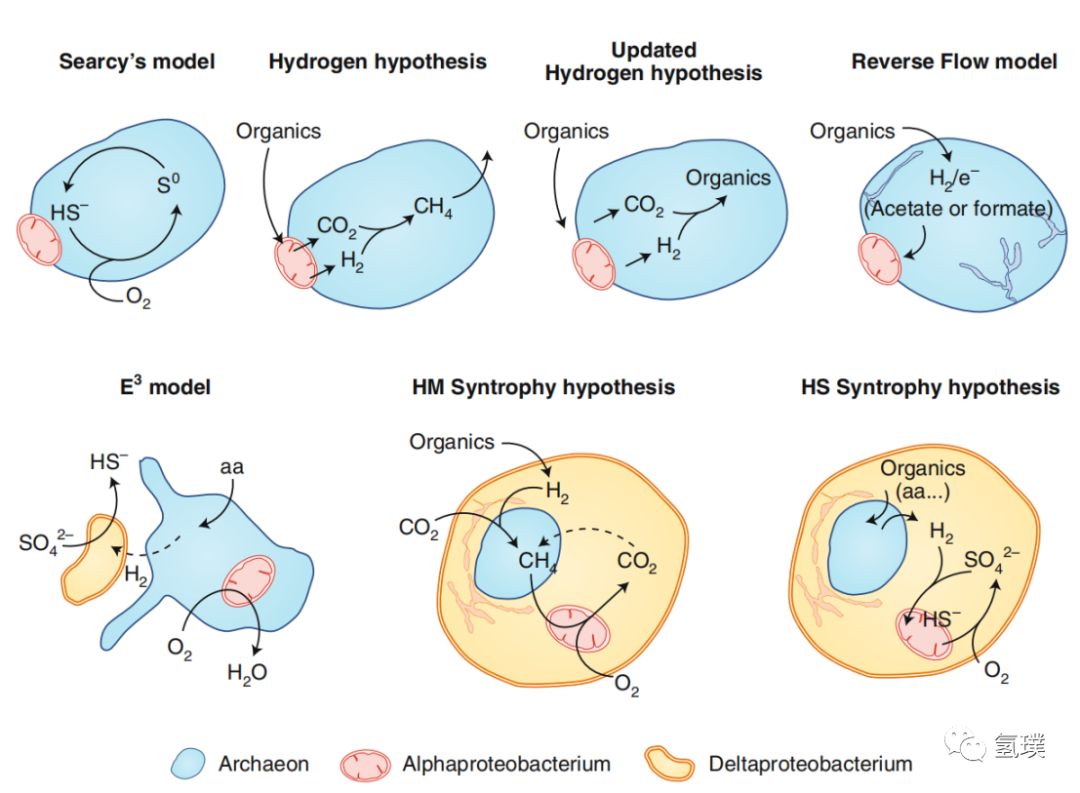
基于代谢交换的真核生物起源共生模型
最初的真核生物进化的氢假说提出了一种氢气介导的共生关系,由分解氢气的产甲烷古细菌和产生氢气的α变形杆菌共生,氢气被用来减少CO2并产生甲烷。在氢假说的一个更新版本中,一种自养的、非产甲烷的古菌代替了最初的产甲烷宿主,它利用线粒体祖先释放的氢来固定碳产生有机物。Purificación等最近提出了基于氢气和硫代谢的真核生物进化的三元共生模型 (HS Syntrophy),考虑了Asgard古菌的发现及其祖先代谢潜力所施加的限制,是比较全面的真核细胞进化模型之一。HS共生模型认为,真核生物起源于元古宙早期的微生物垫,由产氢的Asgard古菌内共生到复杂的硫酸盐还原的δ变形杆菌(sulfide-reducing bacteria,SRB)内,随后线粒体由多功能、兼性需氧、氧化硫化物和无氧光合作用的α变形杆菌(sulfide-oxidizing bacteria,SOB)进一步内共生体进化而来。
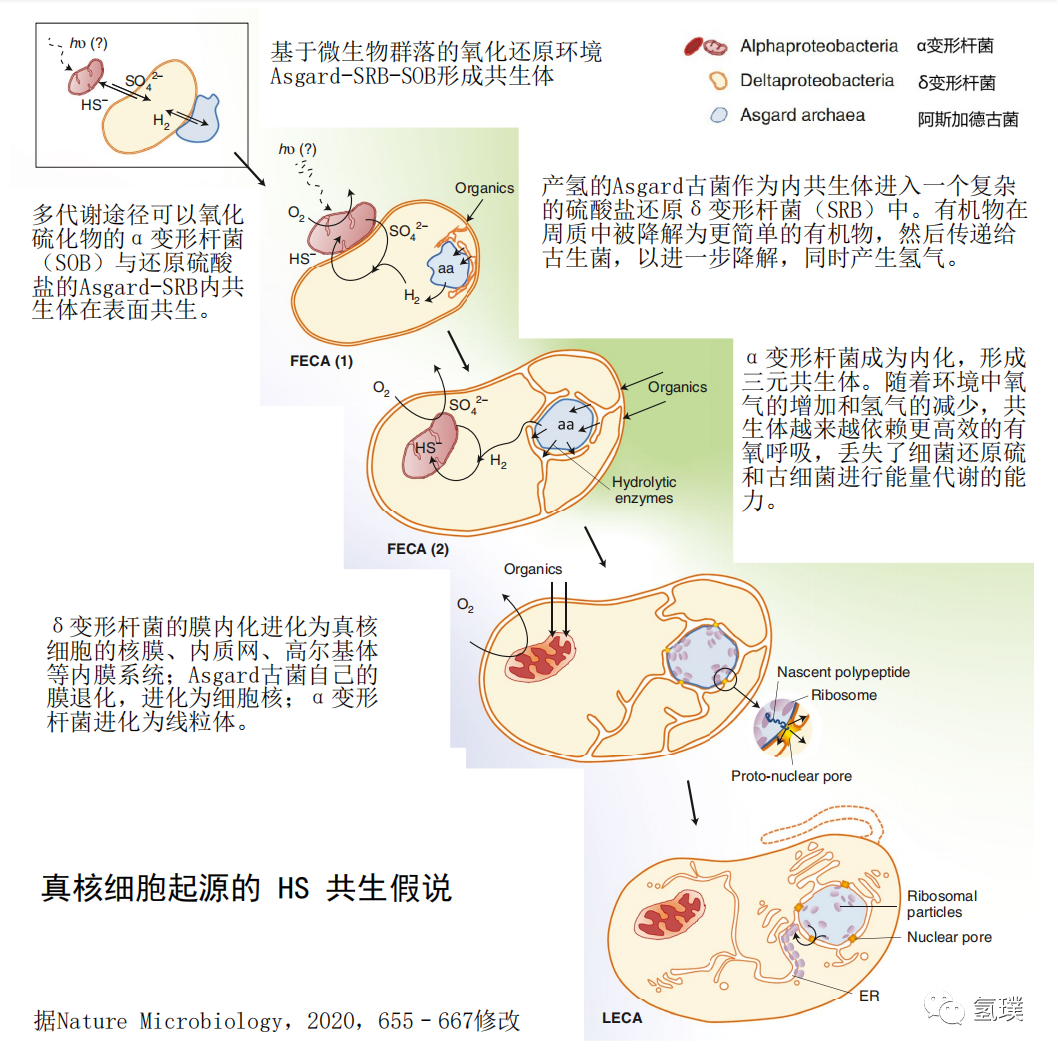
δ变形杆菌的代谢类型多样,可以使用或产生氢或直接产生电子,并经常参与微生物种间的氢或电子转移,其中许多细菌利用硫酸盐氧化有机化合物,但它们也可以是自养的,并可以根据环境条件发酵或在代谢之间进行切换。δ变形杆菌与古细菌建立了广泛的共生关系,与产甲烷菌共生作为供氢者,与甲烷营养古菌共生消耗氢还原硫酸盐。硫还原δ变形杆菌(SRB)还与硫化物氧化或其他细菌、真核生物建立共生关系。这种兼容并包的特性支持 SRB 作为真核生物产生的母体(宿主)。进化过程中真核生物的细胞膜以及逐渐形成的核膜、内质网、高尔基体等庞大的内膜系统也是由δ变形杆菌的膜内化而来,这也意味着如果δ变形杆菌可以使用或产生氢或电子,现在的真细胞核膜系统中可能会保留痕迹,与氢气代谢相关的酶系统可能存在于这些膜的某些分子中,如膜上的氧化还原酶类等生物大分子。
Asgard古菌中包含与真核生物类似的基因,例如洛基古菌,被发现具有真核生物的标志性基因肌动蛋白基因,通过对它们DNA的深入分析,表明所有的现代真核生物来自同一个古菌群。Asgard古菌可以分解有机物生存,并产生氢气作为废气。古菌内共生到 SRB 后,为得到更多的有机物,Asgard古菌逐渐被 SRB 的膜包裹,进化为真核生物的细胞核,古菌自己的膜则退化了。古菌的基因组与真核生物细胞核的基因组有很多共性也就容易理解了。推动Asgard古菌内化的主要动力是 SRB 需要利用氢气还原硫酸盐的代谢需求。
α变形杆菌从代谢类型来看是一个特别多样化的有机体群体,被认为是与线粒体具有最为紧密亲缘关系的细菌物种。在微生物垫的上层,占主导地位依赖 H2S 的厌氧光合细菌与 SRB 相互作用完成硫循环,由此可以推测兼性好氧的线粒体祖先也可能是光合(或混合营养的)细菌。线粒体祖先作为一种必需的内共生体的结合意味着整个内共生体的代谢发生了根本性的变化,由于线粒体祖先是好氧的,并且可以通过直接氧化有机物获得更高的能量,因此共生体开始完全依赖有氧呼吸。这导致了效率较低的厌氧古菌的代谢和细菌硫酸盐还原代谢丢失。同时,原始的真核生物迁移到微生物垫的氧含量高的完全氧化层,在氧化层表面扩散。
氢气在真核细胞进化的三元共生体的形成过程中起到关键作用。随着地球环境氢气来源的减少,宿主就会更加严格地依赖于产氢的共生体,这提供了一种强大的选择力,不可逆转地将共生体和宿主联系起来,如果共生体逃逸,宿主会“饿死”。宿主可以利用自身与环境的表面摄入有机质并提供给共生体,并尽量选择大表面积包围共生体增加接触面,以提供更多的有机质给共生体、获得更多的氢气。氢气还原硫酸盐维持硫循环也是推动线粒体祖先不可逆内共生进入宿主的推动力。随着氧气在地球的逐渐积累,适应环境的进化选择,宿主对氢的代谢依赖性消失了,厌氧环境的限制也消失,宿主成为异养类型,共生体成为现在真核生物细胞核和线粒体等的前体。
真核生物起源的共营养假说提示,真核细胞从富氢的远古地球走来,随着氧气在地球的逐渐积累氢气愈来愈少,Asgard-SRB-SOB 共生体完成了向真核细胞的关键性的几步。通常认为,在新的细胞中,最早存在于共生体古老祖先的产生和利用氢气的氢代谢功能由于缺乏氢气逐渐丢失了。然而,近年来研究揭示氢气对高等生物如动物、植物具有很强的生理功能,我们将不得不重新审视这一观点。
我认为,真核细胞代谢氢气的功能可能并没有丢失,而是留在线粒体、细胞膜、内膜等真核细胞结构中,在现在氧气浓度很高的环境下氢气的代谢被抑制隐藏起来了,合适的条件下氢气代谢依然会发挥作用。另外,虽然空气中的氢气没有了,我们人体却由于肠道菌群等微生物的共生一直沐浴在氢气中,人体的微生物数量甚至比细胞还要多,产氢和用氢的微生物大量存在于肠道菌群,只不过每个个体由于肠道菌群群落结构的不同,氢气的量不一样罢了。
参考文献
1.Martin W , Miklós Müller. The hydrogen hypothesis for the first eukaryote[J]. Nature, 1998,392(6671):37-41.DOI:10.1038/32096
2.Woese, C. R. & Fox, G. E. Phylogenetic structure of the prokaryotic domain: the primary kingdoms. Proc. Natl Acad. Sci. USA,1977,74, 5088–5090. DOI:10.1073/pnas.74.11.5088
3.Laura, Eme, Anja, et al. Archaea and the origin of eukaryotes.[J]. Nature Reviews Microbiology, 2017. DOI:10.1038/nrmicro.2017.133
4.Purificación López-García, Moreira D . The Syntrophy hypothesis for the origin of eukaryotes revisited[J]. Nature Microbiology,2020. DOI:10.1038/s41564-020-0710-4
5.P López-Garia, Moreira D . Metabolic symbiosis at the origin of eukaryotes.[J]. Trends in Biochemical Sciences, 1999, 24(3):88. DOI:10.1016/S0968-0004(98)01342-5






