复合物I极化分解氢气的可能机制
关于线粒体的系列文章,有朋友说专业太强了,不好懂,尽管如此,这些文章的阅读量仍然能过千次,说明总有一些人能看懂,当然也有一些朋友即使不懂也坚持看下去的。为了能让大家对线粒体系列有更好理解,这里我对线粒体进行一些白话解释。
我们的细胞维持功能需要不断消耗能量,细胞能量来自对能量营养素分解产生的化学能。能量营养素就是我们吃的糖脂肪和蛋白质等,这些物质类似汽车的汽油,汽车消耗汽油是用燃烧的方法,但我们的细胞没有汽缸,只有利用酶的催化一步步对这些能量化合物进行分解,一边分解一边把能量变成可使用的化学能小分子ATP。我们可以理解为这些ATP就是电池,可以驱动细胞内的各种工作,例如肌肉运动,神经放电,合成DNA分子等工作。线粒体就是一个ATP电池制造厂,不断提取营养物质的化学能制造这种小电池。这个过程非常神奇,先从能量物质的提取高能级电子,然后利用电子能级的下降经过细胞膜电的驱动合成ATP,虽然ATP仍然是化学能,但线粒体合成ATP过程真是利用的电能。我们将的线粒体复合物I就是一种利用高能电子转移质子,制造细胞化学能的最典型代表。
关键的关键是,这个复合物I和氢气代谢的氢化酶就是一家人,所以这才引起我们的重视。氢气曾经是低等生物的能量转化枢纽,能量转化的分子介质就是氢化酶,这分子能合成氢气,也能分解氢气。作用线粒体复合物I这种氢化酶的后代,代谢氢气的潜力应该仍然存在。今天的故事就是讲,如果线粒体复合物能分解氢气,具体会怎么干?
复合物I氧化还原酶可能从膜结合氢化酶演化来,复合物I的49kD/ PSST亚单位和氢化酶大小亚单位高度同源,两者N2簇、质子传递通路和催化中心周围折叠结构类似,这些是实现能量偶联功能的重要结构。氢化酶和复合物I在结构上同源,但复合物活性中心没有金属离子,氢化酶的金属中心位置被复合物I的85-89位氨基酸(含Tyr87)替补,其功能从分解或合成氢气变成醌结合位点。据推测复合物I祖先分子可能拥有同时使用氢气和醌作为电子和质子载体的能力。高等生物线粒体复合物I也可能仍然拥有祖先特征,就是能同时使用醌和氢气作为电子载体的能力。在这些保守结构中,和催化中心靠近的哺乳动物复合物I的His59或细菌复合物His38最可能是激活氢气的结构。目前对镍铁氢化酶本身的分解氢气共价键精确机制也没有完全阐述,但是研究表明氢气和金属催化剂结合可增加氢气的极性和酸化,然后与附近镍原子结合碱性氨基酸残基如精氨酸质子发生酸碱反应。
分解氢气的机制也可基于无金属氢气激活理论:受阻路易斯酸碱对(FLP)。这种理论模型中,位于路易斯酸碱之间的氢气可被激活,这种酸碱反应被立体空间结构阻止,是酸碱基团虽然接近但距离不足以相互反应。
在[镍铁]氢化酶中碱性残基能参与无金属FLP机制提示,在复合物I中氢气裂解反应也能不需要镍原子作为酸催化剂,而是与亲电反应物发生反应实现。在哺乳动物复合物I中,His59吸收质子碱性氨基酸分布在被氧化N2簇,亲电子/质子的醌自由基能作为反应加速剂,氢气分子激活可提供两个电子和两个质子给电子传递链,这类似复合物I中从醌到氢醌的反应。假定复合物I分解氢气可逐步转移电子,氢气将可代替氢化酶催化中心铁硫簇结合的氢负离子。氢气分子进入复合物I催化中心,这里存在醌形成的亲电子体/亲质子体,包括不稳定半醌中间体,醌头基团能通过牵拉电子激活氢气,类似FLP效应机制分解氢气分子。当亲电子醌提高氢气分子酸性时,位于N2簇附近脱质子碱性基团可激活氢气。Tyr108氧化N2簇能诱导氢气极化,吸收氢气的电子,亲质子醌能吸收氢气的质子。但是现在不知道当半醌自由基存在时,N2簇是氧化态还是还原态。(我的理解:这种FLP效应就是推拉效应,一个酸一个碱,一个亲电子,一个亲质子,这样可把中性的氢气变得具有极性,结果把氢气变成一个氢负离子,一个氢离子,或将氢气的共价键破坏,这比简单从中间切割为两个氢原子容易)。
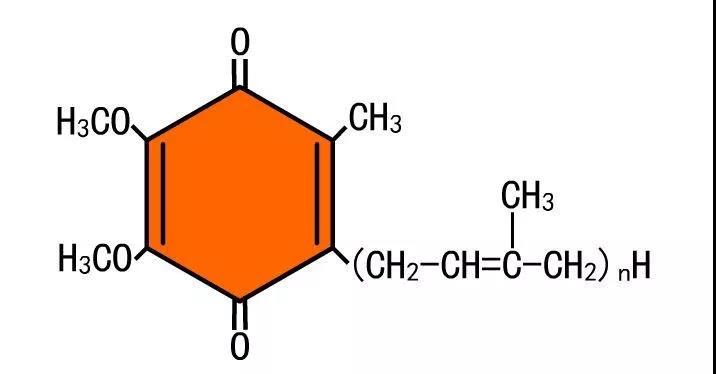
醌是有两个羰基的苯环。
醌不是金属,没有被固定在活性中心,醌头部滑动可产生碰撞能,这是化学反应发生的条件。活性中心结构可塑性和醌头转动能形成一个理想分子内距离,满足氢气发生FLP效应的条件,醌头部和碱性基团和N2簇靠近可共同激活氢气分子。醌和半醌作为亲电子酸,His59作为亲质子碱,亲电子(酸)和亲质子(碱)联合作用产生类FLP效应,将无极性氢气极化激活。类似的情况如在胺氧化酶和乙醇氧化酶活性中心组氨酸羟基就具有亲质子作用。尽管组氨酸不涉及FLP效应,组氨酸的去质子化咪唑基有吸引质子作用,可联合亲电子醌和氧化态N2参与极化氢气。在镍铁氢化酶,参与FLP样效应的精氨酸碱性胍基和镍原子之间的距离在4.5 Å以内,氢分子被局限分解。
在激活态哺乳动物复合物I活性中心,半醌和Tyr108羟基之间的距离小于3 Å,His59残基位于酪氨酸反面。因为醌头部可转动,氢气位于醌头部和His59碱基之间被碾压激活,这也是氢气被FLP样效应激活的理想距离。His55是His59附近另一个保守残基,也可能参与氧化氢气。这种可能性依赖于在复合物I从失活向激活态过渡时醌头部C1或C4基团处于亲电子状态。
靠近活性中心的其他碱性基,可通过FLP样效应从氢气吸收质子,导致这些被囚禁在醌头部周围的氢气被激活。Arg63和Arg53(解脂耶氏酵母是Arg99和Arg89)是候选者,可提供亲质子碱基,但不清楚在激活态变构过程和醌头部的距离是否也符合条件。N2和His190之间距离在4 Å内,His190是氧化还原波尔团,在解脂耶氏酵母是His226,嗜热链球菌是His169。这些结构也具有FLP效应条件,虽然没有醌参与,可能性比较小。
许多复合物I抑制剂属于醌类似物或拮抗物,用抑制剂能研究醌在复合物I激活氢气的作用。虽然在失活态氢气和His59 /N2簇Tyr108靠近还不能在哺乳动物复合物结构中确认,但这种情况非常有可能,考虑到复合物I从铁镍氢化酶进化来,活性中心结构仍保留氢化酶同源的氨基酸残基。重要的是,氢气能缓解缺血再灌注损伤,这符合复合物I失活态减少缺血再灌注过程超氧阴离子的产生,减少复合物I失活激活切换过程的电子泄露。复合物I醌结合位点结构和氢化酶高度同源,他们的活性中心内表面都有许多极性和亲水氨基酸。
在Q室N2簇周围表面因碱性残基表现为正电荷,远处和醌疏水尾结合则为负电荷,这里的结构能阻止其他分子进入醌室。Q室将酸性和碱性部分分离产生静电效应极化Q室内的醌分子,也影响醌头部氧化还原活性和改变进入活性中心醌分子的运动速度。类异戊二烯醌尾进入Q室,氢分子因具有两亲性能进入Q室最底部。Q室内表面保守序列和带电荷残基的任何突变都会破坏复合物I功能并导致疾病发生。
复合物I的Q室基本功能是将电子传递给醌,Q室和氢化酶活性中心同源,根据氢气被分解的酸碱极化理论,结合受阻路易斯酸碱对(FLP)效应,就是在狭小空间内存在靠近但足以发生反应的酸碱基团,这会对非极性的氢气分子产生剧烈的极化,导致氢气被不均匀分裂为氢离子和氢负离子,后者在变成氢离子和两个电子。






