氢化酶和线粒体复合物I的进化关系
氢化酶是细菌代谢氢气的基础,也说明氢气是生物分子的关键证据。利用这种酶,细菌等微生物可以制造和利用氢气,氢气可作为这些微生物的能量介质。氢能源技术中的生物制氢也包括试图能模拟氢化酶的功能,把其他如太阳能转变为氢能。
线粒体是真核细胞发电厂,其最重要的电子传递单元是线粒体复合物,非常有意思的是,线粒体复合物I竟然可能是细菌氢化酶的后代,这不仅证明氢气在生命物质能量代谢过程中的核心地位,也提示氢气在地球生命世界中的潜在广泛作用。本文重点介绍氢化酶和复合物I之间的进化相关性。
氢化酶是线粒体复合物I电子传递功能单元的祖先分子成分,不过受地球大气氧气水平的影响,氢化酶进化为复合物I从分解氢气释放电子演变为传递电子的功能,这是今天生物学的看法。从进化角度看,线粒体复合物I这种源于氢气代谢酶的后代分子,有可能仍然保留着代谢氢气的潜力,相反氢气也可能会对这种反应产生调节作用,这非常符合生物体系中产物就是调节物的普遍现象。因此,线粒体复合物I可能是氢气调节细胞功能的分子靶点,就是我们一种梦寐以求寻找的氢气效应的靶点。从这个角度看,全面了解线粒体复合物和氢化酶的进化历程,对理解其功能,特别是寻找氢气发挥作用的分子靶点,具有重要意义和价值。这正是我最近围绕氢气代谢、氢化酶和线粒体整理相关研究资料的根本目的。
Volbeda A , Fontecilla-Camps J C . The Evolutionary Relationship Between Complex I and [NiFe]-Hydrogenase[M]// A Structural Perspective on Respiratory Complex I. Springer Netherlands, 2012.
摘要:线粒体复合物I全名为NADH泛醌氧化还原酶,含义是这种蛋白分子复合物的功能是氧化还原反应,是将NADH的电子传递给泛醌,使NADH氧化,使泛醌还原的催化酶。
复合物I是有氧生物线粒体内呼吸链的首个元件,在结构上和[镍铁]氢化酶相关。一个比较可信的假说认为,可溶性氢化酶和膜结合逆向转运体联合进化形成复合物I的祖先分子,逆向转运体具有逆向转运质子的功能。研究发现许多微生物都存在这种氢化酶。本文对受环境氧分压选择压力导致氢化酶进化为复合物I的过程进行讨论,通过对复合物亲水区晶体结构和镍铁氢化酶进行比较,推测氧化还原诱导了质子泵4螺旋结构重组。
一、氢化酶和复合物I同源性的发现
Böhm R等1990年代意外发现,线粒体复合物I和镍铁氢化酶存在同源关系。镍铁氢化酶能把氢气氧化和质子跨膜转运偶联起来,这类似线粒体复合物的功能。镍铁氢化酶和复合物I序列类似性最先报道的是大肠杆菌甲酸氢解离酶三个开发阅读框,其中一个和氢化酶大亚单位结构相关。1991年有研究发现,牛线粒体复合物I的75和51kDa亚单位和罗尔斯通菌氢化酶可溶性NAD+氢化酶两个硫酸锌脱氢酶亚单位接近。后来发现复合物I和镍铁氢化酶小亚单位N末端黄素氧化还原蛋白显著类似,多种类似情况见硫酸锌脱氢酶亚单位和去果胚珠型脱硫弧菌氢化酶等。
Böhm R, SauterM, Böck A (1990) Nucleotide sequence and expression of an operon in Escherichiacoli coding for formate hydrogenlyase components. Mol Microbiol 4:231–243
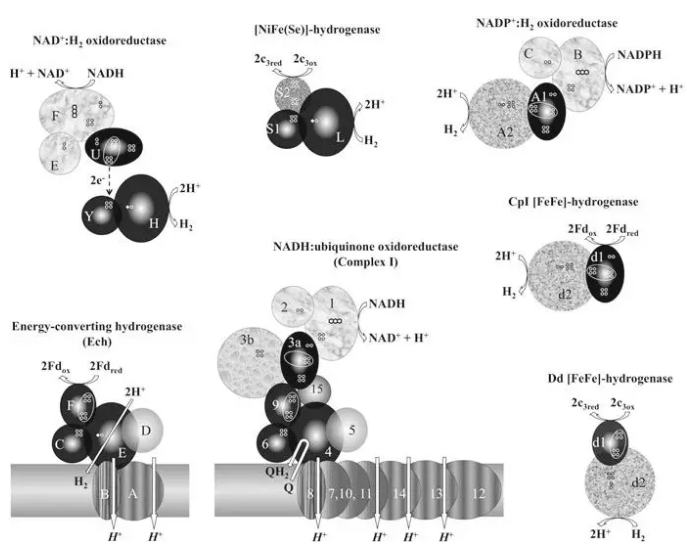
图1复合物I和氢化酶相关的图解。
中间下排是15个Nqo亚单位的完整嗜热菌复合物I模型(Efremov et al. 2010 )。从左顺时针旋转分别是:
巴克菌能量转换氢化酶(Hedderich 2004 )。
集胞藻5同源亚单位组成的NAD(P)+还原镍铁氢化酶(Cournacet al. 2004 )。
异二聚体铁镍(硒)氢化酶(Volbeda et al. 1995 ; Garcin et al. 1999 )。
海栖热袍菌3亚单位NADP+还原铁铁氢化酶(Verhagenet al. 1999 )。
梭菌二型可溶铁铁氢化酶(Peters et al. 1998 )。
去磺弧菌可溶铁铁氢化酶(Nicolet et al. 1999 ).
复合物I和许多氢化酶序列都存在类似性。例如海栖热袍菌三亚基铁铁氢化酶,集胞藻的5亚基氢化酶,膜结合多亚基镍铁氢化酶。膜结合氢化酶有6个保守核心亚基和复合物I相关(Hedderich 2004 )。
这些氢化酶的典型代表如大肠杆菌氢化酶、一氧化碳诱导红螺菌氢化酶和甲烷菌能量转换氢化酶等。和复合物I一样,这些氢化酶的基本功能是质子泵。甲烷菌F 420氢气脱氢酶复合物也具有质子泵功能,其11和13亚单位和复合物I亚单位同源(Deppenmeier 2004 )。不同于用NADH为供氢体,甲烷菌供氢体是F420和氢气,电子受体为1,4-甲桥萘。
二、镍铁氢化酶和复合物I同源的追溯
复合物I和镍铁氢化酶的关系源于能量代谢和大气氧气含量。最早进化步骤是可溶性二聚体镍铁氢化酶和膜结合逆向转运体的聚合,后者主要是K+/H+或Na+/H+逆向转运体(Mathiesen and Hägerhäll 2003 )。
随后是捕获更多氧化还原亚单位,逐渐演化为偶联质子泵的泛醌氧化还原酶。这种复合物氧化氢气,还原泛醌,泵出质子。这种蛋白是细菌复合物I、甲酸脱氢酶、古甲烷菌F420氢气泛醌还原酶和膜结合氢化酶的共同祖先。这些氢化酶小亚基和大亚基和复合物I有高度同源性,这种同源性超过可溶性镍铁氢化酶,这一现象支持这种进化模型(Friedrich and Weiss 1997 )。后期招募逆向转运体和其他亚单位,逐渐失去氢化酶活性,获得/硫辛酰胺脱氢酶活性,变成复合物I。泛醌结合位点居于嗜热链球菌复合物I的nqo4亚单位,和镍铁氢化酶代谢氢气的活性中心拓扑结构接近(图2)。
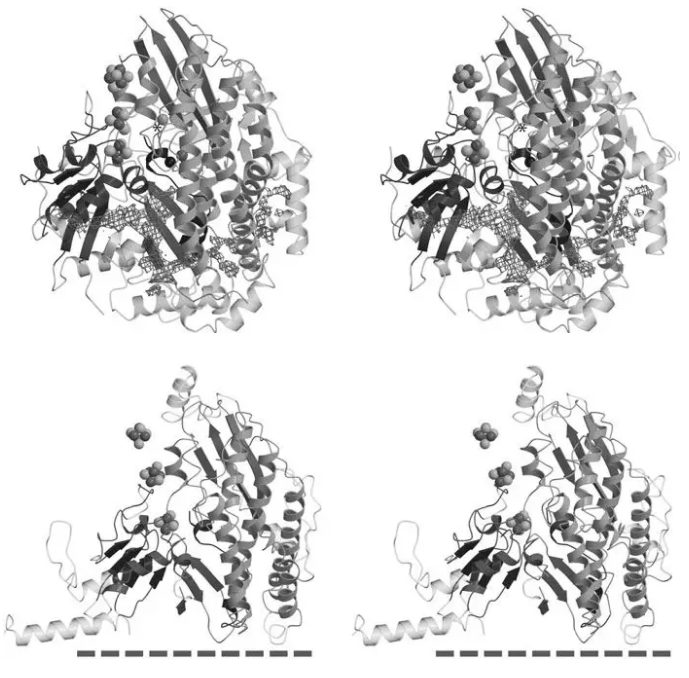
图2嗜热链球菌复合物I的Nqo4和Nqo6亚单位(下)和细菌镍铁氢化酶(上),两者非常神似。细丝网代表气体通道。
大肠杆菌复合物I小膜结合亚单位和膜结合氢化酶镍铁氢化酶大亚单位的谷氨酸和门冬氨酸残基高度同源,这意味着两者的结构和功能相关性。可溶性镍铁氢化酶和膜结合逆向转运体聚集可能是因为许多反应都需要高还原电子。
据推测原始地球大气含有非常丰富的氢气,这解释了可溶解性氢化酶存在的合理性(Tian et al. 2005 )。但是氢气非常容易逃逸到太空,地质时间尺度上地球大气氢气浓度会迅速下降。今天地球大气氢气含量极微量,导致氢气被二氧化碳氧化需要额外能量吸收。有证据表明产甲烷菌二氧化碳氧化氢气的过程需要消耗质子或钠离子电化学梯度能量 (Hedderich 2004 ) 。
偶联氧化还原反应和膜离子梯度热力学反应会导致反向电子传递,或者电子传递和离子梯度之间存在可逆性。能量传递氢化酶的功能是离子泵,和复合物I不同的是,这些酶不用泛醌作为电子受体(Hedderich 2004 ) 。可逆电子传递和二氧化碳还原可能有选择优势形成氢化酶逆向转运体复合物。
复合物I和呼吸链的进化可能和大气氧气浓度提高有关。当地球上光合蓝菌大量繁殖,大气氧气水平逐渐升高,需氧呼吸开始出现。地球大气氧气增加被称为大氧化事件,发生在23亿年前。存在于细菌和古菌体内的细胞色素氧化酶是有氧代谢的关键酶,能把氧气还原为水。如果基因存在水平转移,最后共同祖先(LUCA)在某些情况应使用有氧呼吸。因为LUCA生活在20亿年前,有氧呼吸已经在大氧化事件前进化出。在光合作用出现前,水被光分解产生少量氧气,在氧气浓度比较高的区域进化出需氧呼吸(Kasting 1993 ) 。
在恶劣自然条件下,氧会变成活性更强的单线态氧、超氧阴离子、过氧化氢和羟基自由基等。因为氧气/水的正电位比较高,使用氧气作为电子最终接受体有比较大的进化压力,但也能让低电位氢气释放更多能量。简单说就是,氧气氧化能力比较强大,接受电子可产生更多能量,氧气作为电子接受体在进化上有更多优势,虽然活性氧也会给生物体带来较大应激。
马丁和米勒提出真核线粒体进化的氢假说。氢假说认为,真核细胞来自一种古菌和一种变形菌共生,变形菌给古菌提供氢气(Martin and Mueller 1998 ),古菌成为真核细胞主体,变形菌进化为细胞内线粒体。这种观点存在争议,因为没有遗传学证据证明存在能产生氢气的变形菌。也有人认为真核细胞来自代谢氢气的古菌和一种需氧呼吸变形菌。许多研究证据表明,变形菌是线粒体的祖先,借助有氧呼吸,变形菌消耗氧气能减少氧气的毒性,这让古菌获得了高氧条件下的生存优势。
三、氢化酶和线粒体复合物I的结构关系
嗜热链球菌复合物I的3.3 Å分辨率晶体结构显示其Nqo3、Nqo4、Nqo6和Nqo9亚单位三维结构类似于镍铁氢化酶和铁铁氢化酶(表2 , 图1),其氨基酸序列也有同源性。如图1所示,在FMN和Nqo1黄递酶中心和Nqo4泛醌还原位点之间的电子传递路线上有7个铁硫簇。靠近FMN的Nqo2内末端双铁硫簇功能是抗氧化剂。Nqo3C末端铁硫簇位置太偏无法参与电子传递。根据和铁铁氢化酶序列类似,推测一些复合物I可能也参与外部氧化还原亚基电子传递过程。
2010年解析的复合物I有7个疏水膜结合亚单位,其中Nqo12和Nqo8分别和甲烷八叠球菌能量转换膜结合氢化酶亚单位A和B同源。Nqo12和Nqo8也和细菌钠或钾逆向转运体同源,这符合质子泵的特征。另外Nqo12、Nqo13和 Nqo14也有同源性。
复合物I和NAD+还原镍铁氢化酶HoxEFUYH类似。按照氨基酸序列,这种酶应包含2个铁硫簇,其中1个在HoxU和第二个4铁硫簇相关,后者在铁氧化还原蛋白样亚基Nqo3和Nqo9内。铁氧化还原蛋白样亚基能把黄递酶和氢化酶序列连接起来。在NADP+还原铁铁氢化酶中,同样具有连接黄递酶和H簇域的作用,也和简单可溶性氢化酶类似。复合物I泛醌结合亚单位Nqo4和4铁硫簇亚单位Nqo6分别高度类似于镍铁氢化酶大亚基和小亚基N末端。
尽管这些酶催化的反应有很大差别,但序列和Nqo4和Nqo6大多数二级结构同源性非常高。镍铁氢化酶有大小亚基之间疏水气体通道,是活性位点和分子表面联系通路。这个气体通道在复合物I变成一个更宽大通道,面向复合物I膜表面(图2)。复合物I这一区域可能变成了泛醌结合位点。Albracht等报道,牛复合物I有两个不稳定FMN,根据结构和镍铁氢化酶类似推测结合在Nqo4亚单位。后来发现,这种不稳定FMN也存在于罗尔斯顿氏菌可溶性镍铁氢化酶。这种不稳定FMN的发现对理解复合物I的功能有重要意义。这意味着对这些分子纯化后必须保持双功能酶活性才符合自然状态。Nqo4和Nqo6之间大部分朝向膜表面有利于提高和醌分子和FMN的结合机会。但是人们对这个空穴的精确形态大小还不清楚,因这些蛋白这个区域仍没有被解析。采用结构叠加方法,发现FMN不能和黄素意义结合,因这会导致其他结构的塌陷。对Nqo4和Nqo6与镍铁氢化酶序列比较发现存在许多保守氨基酸序列。这些氨基酸分布在整个肽链上,主要分布在催化活性中心和近端铁硫簇。大多数13个甘氨酸位于β折叠和α/310螺旋前后,是维持主链构象的重要因素。许多保守疏水氨基酸位于疏水内区。4铁硫簇4个半胱氨酸配位中3个在Nqo6亚单位是保守序列,最接近N末端的是醌结合通道。另外大量亲水氨基酸残基属于保守序列,大部分都深埋于蛋白质结构内部。比较有吸引人的推测是,这些氨基酸和质子传递有关。复合物I两个电子还原泛醌伴随两个质子跨膜转运。复合物I和镍铁氢化酶可能有类似的质子转运通路。虽然复合物I3.1 Å分辨率亲水结构没有包括水分子,在保守的Asp401和Asp139之间存在一个亲水洞穴。氢化酶没有这种洞穴,相关空间被3个保守的组氨酸占据。Asp401相关天冬氨酸残基和镍铁氢化酶质子转运功能相关,面对膜表面Asp139和质子跨膜转运相互作用。在复合物I和能量转化氢化酶质子可通过和一个链接膜连接的通路进行转运。为避免质子在转运路途泄露,这些通路不能被同时激活。其功能可能通过氧化还原状态调节,然后启动蛋白结构的变化。镍铁氢化酶大亚基长四螺旋束有3个α螺旋。这些螺旋变成一个长结构分隔为两个区域,在Asp528 和Asp129之间产生一个亲水洞穴。这和复合物I中Asp401 和Asp139之间的结构情况类似,有利于质子跨膜运输。

图3 复合物I的Nqo4亚单位和铁镍氢化酶大亚单位(Df和Db)的结构对比。
有些功能可能需要区域移动实现。例如镍铁氢化酶大单位的不变His219和Arg63通过水网和小亚基C末端和黄素样区近端连接。在复合物I前者被铁氧化还原样亚单位代替。复合物I内的4铁硫簇氧化还原电位具有pH依赖性,可被组氨酸169调节。如果突变蛋氨酸能消除pH依赖性,但对质子泵的效率没有影响。因此质子泵不依赖于这些铁硫簇的氧化还原电位,改变复合物I的氧化还原电位和泛醌的质子化状态或许可以控制质子流动。类似机制可能也操纵氢化酶的镍铁离子活性。
四、结论
比较强的证据表明,线粒体复合物I是从细菌镍铁氢化酶在进化来。能量转化氢化酶是进化的第一步,这种结构是可溶性氢化酶和质子泵结合产生。许多证据表明,有氧呼吸在大氧化事件前进化产生。值得注意的是,复合物I和镍铁氢化酶都是真核细胞进化产生的主要参与者。






